
by Gertrud U. Rey
Herpes simplex viruses infect cells of the skin and mucous membranes, where they establish a lifelong persistent infection in sensory neurons. Sporadic reactivation and viral shedding may lead to painful oral and genital disease and a three to five-fold increased risk of HIV transmission. There is currently no vaccine to prevent infection with herpes simplex virus type 1 or type 2 (HSV-1 or HSV-2).
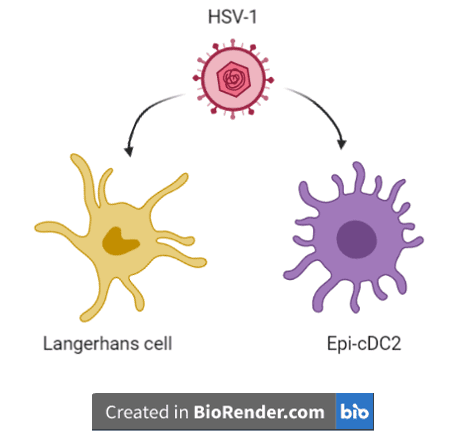
Until recently it was thought that initial interactions of HSV-1 with the immune system only involve Langerhans cells. Langerhans cells are skin-resident sentinel macrophages that detect microbial antigens, and they engulf, process, and present these antigens to T cells for downstream immune functions. However, a recent study suggests that early during infection, HSV-1 also interacts with a newly identified immune cell known as an epidermal conventional dendritic cell type 2 (Epi-cDC2). Like Langerhans cells, dendritic cells can swallow microbe-infected cells and present the microbial antigens to T helper cells, ultimately triggering the actions of cytotoxic T cells, which directly kill infected cells.
The study aimed to better define the role of Epi-cDC2s in early HSV-1 infection using ex vivo explants as a model system. The explants consisted of pieces of human inner foreskin that were mounted on specialized gelatin scaffolds to mimic the in vivo environment encountered by HSV-1 during infection. The authors exposed the explants to an HSV-1 virus in which a viral membrane protein was fused to a green fluorescent protein (GFP), allowing them to visually track a resulting infection using a fluorescence microscope. This method revealed that at 24 hours after exposure to the GFP-tagged HSV-1, both Langerhans cells and Epi-cDC2s contained the virus in their cytoplasm, suggesting that these cells either engulfed HSV-1-infected skin cells and/or were themselves infected by HSV-1.
To determine whether the presence of HSV-1 in the cytoplasm of Epi-cDC2s resulted from infection and replication and not just from engulfing infected skin cells, the authors first did the following. They exposed cell cultures of Epi-cDC2s to HSV-1. After six hours of this exposure, Epi-cDC2s contained about as much virus as did control Langerhans cells, which are known to be infected by HSV-1. However, at 18 hours, Epi-cDC2s contained significantly higher HSV-1 levels than Langerhans cells, suggesting increased entry/uptake of virus into Epi-cDC2s compared to Langerhans cells. Next, to assess whether HSV-1 was also replicating in the Epi-cDC2 cells, not just entering them, the authors treated the cells with a fluorescent antibody that binds ICP27, a viral protein needed for replication. A significantly greater portion of Epi-cDC2s than Langerhans cells expressed ICP27, suggesting that HSV-1 was replicating, and doing so more efficiently in Epi-cDC2s.
Viruses may enter a host cell by a variety of mechanisms. One common mechanism, called receptor-mediated endocytosis, involves the formation of cell membrane-derived vesicles. In one version of this process, which requires a low pH, viral binding to a cell surface receptor triggers the cellular membrane to fold inward and form a slightly acidic €œendosome€ around the virus. Another version of receptor-mediated endocytosis is not dependent on a low pH, but requires cholesterol molecules and the motor protein actin to form cell surface protrusions called “ruffles.” When the ruffles become large enough, they collapse back onto the membrane and form large fluid-filled vesicles encasing the virus. In both of these versions of receptor-mediated endocytosis, the resulting vesicles enter the cytoplasm, where they eventually release their contents. In yet another mechanism of entry, also independent of acidic pH, viruses may simply fuse with the plasma membrane and deliver their contents into the cytoplasm.
Although HSV-1 can enter cells by any of these pathways, its entry mechanism differs in different types of cells. To determine which pathway HSV-1 uses to enter Langerhans cells and Epi-cDC2s, the authors treated both types of cells with a drug that prevents acidification of endosomes. They then infected the cells with the GFP-tagged HSV-1 and measured infection by quantitating GFP with a fluorescence microscope. Increasing doses of the drug led to increased inhibition of infection of Langerhans cells, suggesting that these cells are infected with HSV-1 via a pH-dependent mechanism. In contrast, the drug did not affect infection of Epi-cDC2s, suggesting that HSV-1 does not require an acidic pH for entering Epi-cDC2s.
To determine whether HSV-1 entry into Epi-cDC2s occurred via actin and cholesterol-dependent endocytosis, the authors treated Epi-cDC2s with inhibitors of actin or cholesterol prior to infection. Both treatments led to significant reduction in GFP fluorescence inside the cells, suggesting that cholesterol and actin are both important mediators of HSV-1 entry into Epi-cDC2s.
Langerhans cells express a cell surface receptor called langerin, which mediates entry of HIV and influenza A. To see whether this receptor is also required for entry of HSV-1, the authors infected Langerhans cells with HSV-1 in the presence of an antibody that neutralizes langerin. This inhibition of langerin expression led to diminished infection of Langerhans cells, suggesting that langerin is required for HSV-1 entry into them. In contrast, inhibition of langerin on Epi-cDC2s had no effect on HSV-1 infection efficiency, suggesting that, even though Epi-cDC2s do express some langerin, this receptor is not required for HSV-1 entry of these cells.
HSV-1 and HSV-2 are of high public health concern, and a vaccine to prevent infection with these viruses is urgently needed. Immune control of HSV-1/-2 infection and resolution of genital herpes lesions requires the collective action of various types of T cells, which are likely primed by different dendritic cell subsets. Understanding the dynamics of the initial interactions of HSV-1 and HSV-2 with cells of the immune system may result in better strategies for HSV-1/-2 vaccines. The pathways described here have important implications in vaccine design and prevention of persistent infection of neuronal cells.
[The material in this blog post is also covered in this video.]